Рабочее Совещание
"Сверхслабые воздействия на физико-химические и биологические системы. Связь с солнечной и геомагнитной активностью"
6-8 мая 2002 года, Крымская Астрофизическая обсерватория НАН Украины.
Л.В.Белоусов
Взаимодействие биологических и небиологических источников сверхслабого
излучения.
(Стенограмма доклада)
Прежде всего я хотел бы поблагодарить за приглашение на эту замечательную конференцию. На меня, как и на нас на всех, производит глубокое впечатление обстановка этой конференции, поистине настраивающая на связи с космосом. Проблема, которую мы будем сегодня обсуждать – это действительно комплексная синтетическая проблема, которую именно в таких местах и в такой атмосфере нужно обсуждать и искать решение.
У нас, у биологов, занимающихся проблемой сверхслабых излучений, создается впечатление, что мы находимся не просто на стыке классических наук, в их пограничной зоне, но и невольно принимаем участие в создании какой-то синтетической науки, определенное название которой пока сложно дать. То, что о чем здесь будет идти речь несомненно относится не только к биологии, может быть не только к физике, а к некоторым наиболее глубоким взаимодействиям, которые существуют между естественными объектами любой природы.
Я буду в основном говорить о взаимодействии биологических объектов через посредство сверхслабых излучений, но, кроме того, немного коснусь, если позволит время, взаимодействия биологических и небиологических объектов именно в этом смысле.
Мне хотелось бы напомнить, что исследования в области сверхслабых излучений начались на вот этой самой земле в Таврическом Университете, представители которого здесь присутствуют.
Ровно 80 лет тому назад профессор Таврического университета Александр Гаврилович Гурвич, поставил классический опыт по осуществлению оптического, только оптического контакта между двумя корешками лука. В результате после некоторой экспозиции было обнаружено достоверное повышение числа клеточных делений на той стороне вертикального корешка, на который «смотрел» корешок горизонтальный (обзоры: Rahn, 1936; Гурвич и Гурвич, 1945).
Было поставлено несколько десятков таких опытов. В подавляющем большинстве из них результаты заведомо выходили за пределы ошибок, то есть наблюдали достоверное увеличение числа клеточных делений. Я это говорю для того, чтобы подчеркнуть, что с самого начала тематика сверхслабых излучений была связана со взаимодействием.
Первым опытом в этой области был опыт по взаимодействию двух биологических объектов. В дальнейшем эта направленность сохранялась, однако, до сих пор не создано моделей, удовлетворительно описывающих это взаимодействие.
Ясно, что мы здесь имеем дело с очень сильным нелинейным, неаддитивным миром, к которому не применимы относительно простые понятия, скажем такие как сложение излучений, или классический резонанс. В этом мире связи и явления гораздо более глубокие.
В этом докладе я хотел бы остановиться на некоторых конкретных фактах, не делая попытки создать модель или более или менее законченную теорию этого взаимодействия.
Вторым опытом, о котором я хотел бы здесь напомнить, был опыт или серия опытов, поставленные французским исследователем Магру (Magrou) в 30-е годы (см. Гурвич и Гурвич, 1945). Речь идет, опять таки, об опытах по оптическим взаимодействиям объектов через посредство сверхслабых взаимодействий без всяких иных контактов.
Магру использовал два герметически закрытых сосуда, верхний из которых имел кварцевое дно, и был поставлен на нижний. На дне верхнего сосуда лежали развивающиеся яйца морского ежа, в нижнем сосуде размещались источники сверхслабых излучений – это была, как правило, культура бактерий.
Статистически достоверно было показано, что такие взаимодействия приводят к угнетению развития зародышей морского ежа и к возникновению всевозможных аномалий. В свое время эти работы произвели достаточно большой шум, но потом они были забыты, поскольку не находилось убедительных объяснений полученных результатов.
И вот совсем уже недавно мой коллега по Московскому Университету Александр Борисович Бурлаков провел очень большую серию опытов на зародышах рыбы-вьюна, используя примерно ту же установку, что и Магру , хотя о его опытах тогда не знал. А.Б. Бурлаков получил и продолжает получать очень ясные и статистически достоверные результаты (Бурлаков с соавт. 1999а,б; 2000). Полученные результаты сводятся к следующему. Приводятся в оптический контакт зародыши близких стадий развития или зародыши дальних стадий. Если приводятся в контакт зародыши близких стадий развития – например, самых ранних стадий 0 и 1 – то развитие зародышей стадии 0 несколько ускоряется. Т.е. если стадии очень близкие, то развитие как бы подстраивается одно под другое и ранняя стадия ускоряет свое развитие. Однако, если разница между стадиями достаточно велика, то эффект обратный: ранние стадии отстают в развитии. Кроме того, было показано, что смертность на зародышей более ранних стадий достоверно увеличивается, а если стадии близкие, то этого эффекта нет.
Эти эффекты передаются через кварц и не передаются через стекло, т.е. так же как в более старых опытах, начиная с опытов Гурвича.
Дальнейшая задача состояла в том, чтобы понять, а что же, собственно, здесь действует, что именно исходит от зародышей более поздних стадий развития такое, что тормозит, угнетает и подавляет развитие более ранних стадий развития. Заметим, что обратные влияния тоже наблюдаются.
При более детальном изучении обнаружился ряд загадок.
Первая из них заключается
в том, что если вы просто будете измерять интенсивности излучения, то не найдете сколько-нибудь существенных различий по стадиям, скорее даже с увеличением стадий интенсивность излучений ослабляется , но все находится это на уровне фона. И вопрос о «действующем начале» остается открытым.
Поскольку «лобовая атака» закончилась неудачей, решили применить анализ Фурье.
Если рассмотреть спектры Фурье в диапазонах периодов от долей секунды до несколько десятков секунд, то можно увидеть следующее (рис.1).
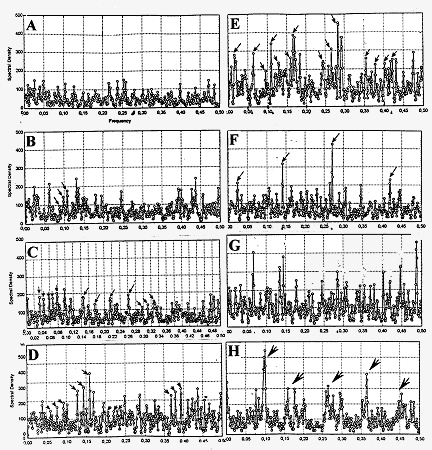
Рис 1Спектры Фурье сверхслабых излучений контрольных образцов (А-D) и развивающейся икры вьюна (Е-Н). А: темновой фон ФЭУ; В: кварцевая кювета с водой; С: неоплодотворенные яйца; D: уродливые нежизнеспособные зародыши. Е: дробящиеся яйцеклетки; F,G: развивающиеся зародыши более поздних стадий; Н: свободноплавающие личинки 4 дней. Горизонтальная ось: частоты, вся шкала соответствует 1 Hz. Вертикальная ось: спектральные плотности. Стрелки указывают на главные максимумы спектральной плотности.
Три диаграммы из левого столбца показывают спектры Фурье контрольных образцов. Здесь просто зародышей нет, здесь вода. Это тоже кювета с водой, это – неоплодотворенные яйца, а вот здесь вот внизу аномально развивающиеся зародыши.
И вот если сравнить эти колонки со спектрами Фурье развивающихся зародышей разных стадий развития, то оказывается, что набор частот более сильно выражен , чем в левой контрольной колонке. Первый вывод, который мы должны сделать – это что зародыши действительно обладают определенным набором частот и если что и действует, так вот именно этот набор частот . Надо сказать, что эти наборы частот иногда составляют гармоники, что само по себе, наверное, и неудивительно.
Следующий вопрос состоит в том, будут ли видны некие достоверные различия по стадиям между спектрами Фурье.
Оказывается удобным представлять спектр в виде автокорреляций – рассматривается автокоррелограмма целого спектра, при этом картина спектра несколько загрубляется, но зато можно «вытащить» некоторые значимые его характеристики.
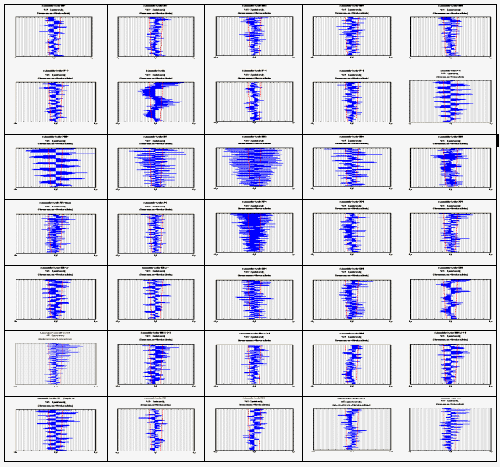 Рис 2 Автокорреляционная структура спектров Фурье на последовательных 2-минутных отрезках времени. Строчки сверху:
Рис 2 Автокорреляционная структура спектров Фурье на последовательных 2-минутных отрезках времени. Строчки сверху:
1 - кювета с водой. 2 - неоплодотворенные яйцеклетки. 3 - 70 мин после оплодотворения (начало 1-го деления дробления). 4: стадия 10 (поздняя бластула) 5: стадия 11-12 (начало эпиболии) 6: стадия 18-19 (формирование органов). 7: предличинки 2,5 дней после вылупления. Косые стрелки указывают на присутствие «змеевидных» фигур
Если мы составим автокоррелограммы спектров разных стадий развития (рис. 2) – вот здесь неоплодотворенная яйцеклетка, а вот здесь через 1 час после начала развития эмбриональная стадия развития, здесь уже личинки продвинутые, то увидим, что по автокоррелограммам имеются весьма четкие стадийные различия. Чтобы понять, что это означает, нужно еще раз рассмотреть спектры. Ясно видно, что на стадиях сразу после оплодотворения наблюдаются очень четкие периоды, которых нет на неоплодотворенных яйцеклетках , а на более поздних стадиях наблюдаются другие наборы частот.
Т.е. действительно, некоторые частотные и спектральные характеристики меняются от одной стадии к другой и, значит, можно думать, что именно эти частотные характеристики оказываются ответственными за эффекты, наблюдавшиеся в опытах Бурлакова.
Теперь дальше мне вот о чем хотелось бы сказать. Результаты взаимодействия можно разделить по времени на две фазы. Первая фаза – это то, что происходит немедленно после начала оптического взаимодействия между объектами. Вторая фаза – это то, что происходит через длительный промежуток времени, через 12 – 24 часа, когда уже наблюдаются морфологические следствия оптических взаимодействий.
Наблюдение за результатами кратковременного взаимодействия осуществляется следующим образом.
В нашем распоряжении имеется большая темная камера с фотоумножителем, катод которого поглощает фотоны. В камеру одновременно ставятся две кюветы, заполненные зародышами.
Когда нас интересует, что происходит, скажем, в течение одного часа после начала взаимодействия, то кюветы ставятся так, что они «видят» друг друга и записывается совокупный сигнал, учитывающий их взаимодействие. Потом между кюветами ставим черную бумагу и записываем изменение излучения, когда они «не видят» друг друга.
Были получены стабильные, четко повторяющиеся результаты.
Вопрос: Какая величина измерялась для (неразборчиво) …..
Ответ: Измерялся поток фотонов в диапазоне от 300 до 800 нм. Период накопления был в основном порядка 0,1секунды.
Ультрафиолет и видимый свет вместе.
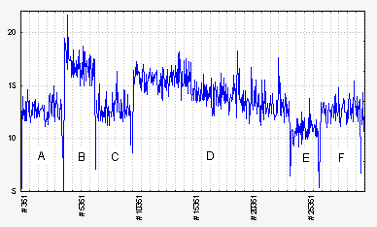
Рис 3 Изменение интенсивности сверхслабого излучения двух разновозрастных партий икры вьюна при их оптическом взаимодействии. А: излучение от партии икры на стадии 2 бластомеров. В: совместное излучение этой стадии + порция икры на значительно более поздней стадии (№ 19-20). С: вторая порция икры убрана, снова светят только 2 бластомера, уровень их излучения пока не изменился. D: вновь светят обе порции вместе – уровень излучения плавно понижается. Е: только 2 бластомера – уровень ниже, нежели на А, С.
F: снова обе порции икры вместе. Уровень значительно ниже, чем на В. Вывод – понижение интенсивности излучения при оптическом взаимодействии. Горизонтальная ось: время секундах. Вертикальная ось: количество импульсов в секунду
Рассмотрим следующий рисунок (Рис. 3). Сначала мы ставим кювету с зародышами очень ранней стадии развития – 2 бластомера. Назовем ее стадия 1. Получаем некий сигнал (Рис. 3А). Потом подсаживаем кювету с икрой другой, более поздней стадией (Рис. 3В). Пусть это будет стадия 2. Сигнал вырос, что вполне естественно. Однако он быстро начинает падать, хотя при этом внутри установки ничего не меняется. Затем убираем стадию 2 (Рис. 3С) – сигнал возвращается к прежнему значению. Возвращаем стадию 2 и держим ее долго (Рис. 3D), то есть начинаем длительное оптическое взаимодействие между икрой двух разных стадий. Теперь мы наблюдаем сильное падение их совокупного сигнала, после чего даже если убрать стадию 2, то стадия 1 светит меньше своего исходного значения (Рис. 3Е).
Основной вывод из этих опытов (которых было много) такой. Когда мы ставим два образца вместе, то излучение сильно нестационарно, вначале оно высокое, а потом оно плавно падает. Такой результат повторяется много раз. Вот, скажем, здесь стоит одна стадия, светит так, потом мы подставляем вторую стадию, сначала она начинает светить больше, но затем сигнал падает.
Потом, когда они перестают видеть друг друга, то сигнал выходит на стационарный уровень.
Статистика была «наведена» таким образом.
Оказалось удобнее всего мерить количество нулей, поскольку мы ведем измерения за промежутки времени в 0,1 секунды. За какое количество периодов у нас вообще нет регистрации фотонов? Какой у нас процент нулевых значений за весь период регистрации? Тогда получается следующее (рис. 4):
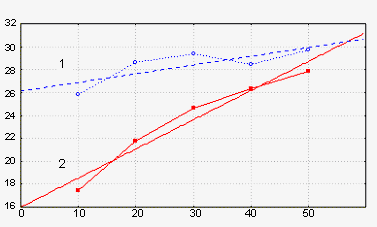
Рис 4. Изменение процента нулевых значений сигналов в течение первых 50 минут наличия (кривая 2) или отсутствия (кривая 1) оптических взаимодействий двух групп зародышей, находящихся на стадиях 6 и 11. Абсцисса – минуты. Ордината – процент нулевых значений.
Здесь у нас две линейно аппроксимированные экспериментальные кривые. Вот одна кривая – кривая 1 - стоят две кюветы с зародышами и они не «видят» друг друга и количество нулей у них за время экспозиции лишь слабо возрастает.
Теперь те же зародыши тех же стадий развития, но они начинают «видеть» друг друга и оказывается, что количество нулей начинает расти и за время экспозиции возрастает вдвое. То есть линеаризованные кривые достоверно различаются по тангенсам своих наклонов: в случаях оптического взаимодействия они выше (Табл. 1).
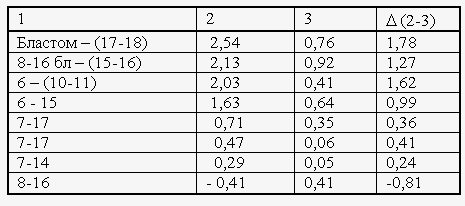
Табл. 1. Динамика нулевых значений сверхслабых излучений в течение первого часа совместной инкубации двух оптически взаимодействующих (столбец 2) и невзаимодействующих, но совместно инкубируемых (столбец 3) групп яйцеклеток или зародышей вьюна. Столбец 1 – стадии развития обеих групп. В таблице приводятся средние значения наклонов (тангенсов углов х 104) линеаризованных графиков нулевых значений, измеренных в последовательные 10-минутные отрезки времени
Вопрос: Что такое количество нулей?
Ответ: Количество нулей – это процент тех периодов накопления фотонов, когда сигнала нет совсем. Опыт ставится так: мы делаем 10 минутные измерения, а период накопления фотонов 0.1 сек - 6000 измерений. За эти 6000 измерений мы смотрим в скольких процентах от 6000 измерений у нас результат нулевой.
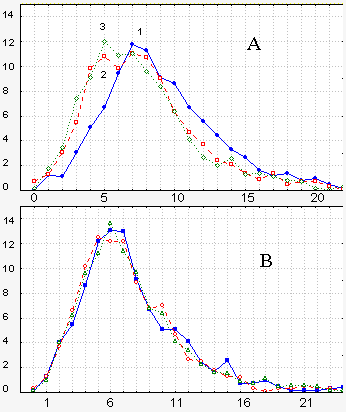 Рис 5Зависимость кривых распределения интенсивностей сигналов от наличия или отсутствия оптических взаимодействий между группами зародышей, находящихся на начале дробления и на стадии 34. Кривые относятся к суммарным сигналам от обеих групп за 10-минутные промежутки времени. Периоды накопления фотонов 0,5 с. .А: кривые 1, 2, 3 соответствуют 10, 20 и 30 мин оптических взаимодействий. Видно смещение максимальных значений в область меньших величин. Б: группы оптически не взаимодействуют (разгорожены черным экраном). Приведенные кривые, записанные через 100 мин одна после другой, практически совпадают. Абсциссы – количество импульсов за период накопления. Ординаты – проценты встречаемости данных сигналов
Рис 5Зависимость кривых распределения интенсивностей сигналов от наличия или отсутствия оптических взаимодействий между группами зародышей, находящихся на начале дробления и на стадии 34. Кривые относятся к суммарным сигналам от обеих групп за 10-минутные промежутки времени. Периоды накопления фотонов 0,5 с. .А: кривые 1, 2, 3 соответствуют 10, 20 и 30 мин оптических взаимодействий. Видно смещение максимальных значений в область меньших величин. Б: группы оптически не взаимодействуют (разгорожены черным экраном). Приведенные кривые, записанные через 100 мин одна после другой, практически совпадают. Абсциссы – количество импульсов за период накопления. Ординаты – проценты встречаемости данных сигналов
Обратимся к рис. 5. Здесь взаимодействуют зародыши двух стадий. Мы ставим вместе и измеряем, составляем кривые распределения в течение разных промежутков времени. Здесь период накопления фотонов больше, чем в прежних опытах (около 1с), поэтому здесь нулей меньше. Но в принципе получается то же самое. Вот мы ставим их взаимодействовать, вот у нас такая кривая распределения и потом по мере взаимодействия максимум смещается влево, т.е. в сторону меньших значений. Это если зародыши «видят» друг друга, т.е. если они не разгорожены черным экраном. Если же зародыши «не видят» друг друга, то это все как бы стоит на месте, количество значений не меняется.
Т.е. первый вывод, который можно сделать, состоит в том, что в самом начале взаимодействия, если зародыши «видят» друг друга, их суммарный сигнал плавно падает или, иными словами, количество нулевых значений плавно возрастает.
Я не буду говорить о попытках объяснения этого эффекта потому, что моя основная цель представить некоторые данные, но если будут вопросы, я постараюсь на них ответить.
Другой критерий, который может помочь при оценке этих взаимодействий – это оценка так называемого куртозиса. Куртозис – это функция четвертой степени момента, А.А. Конрадов может сказать точнее, куртозис – это такой общепринятый показатель количества «пиковости», количества и высоты пиков вот в данной записи.
Надо сказать, что запись сверхслабых излучений, которые выполнены при малых временах накопления фотонов, имеет характер набора вот таких вот пиков.
Именно набор этих пиков всегда один и тот же и для контрольных, и для опытных образцов.
Так вот, оказалось, что если взаимодействуют зародыши с сильно раздвинутыми стадиями развития, разница между которыми составляет 15-17 номеров, то у более ранних зародышей куртозис после взаимодействия падает, т.е. количество пиков уменьшается, а у более взрослых зародышей количество пиков увеличивается, идет некий обмен пиками. Если разница между стадиями меньше, то результат менее определенный, а если разница еще меньше, то рост числа пиков может наблюдаться и в ранней, и в более поздней стадии (Рис. 6).
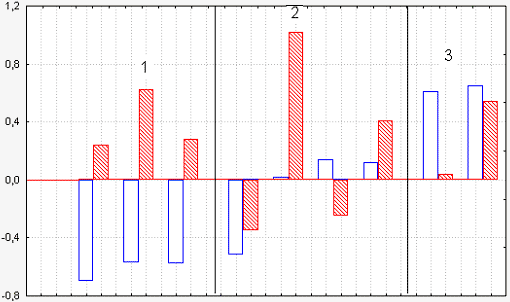
Рис 6 Сводная диаграмма изменений куртозиса после 1-часовых оптических взаимодействий групп зародышей с разными межстадийными интервалами. 1 – интервал 15-17 стадий; 2 – интервал 9-10 стадий; 3 – интервал 4-5 стадий. Заштрихованные столбцы – изменения куртозиса старших партнеров, светлые столбцы – младших партнеров. Ордината – десятые доли. Видно, что при наибольшем разрыве в стадиях развития (группа 1) в старшей группе куртозис увеличивается, а в младшей – убывает
Покажу еще один результат (рис. 7). Это по спектрам Фурье. Одна партия зародышей до начала взаимодействия, мы пишем ее спектр Фурье. Он получается такой . Мы берем вторую партию, пишем до начала взаимодействия ее спектр. Он получается другой. Теперь мы приводим их во взаимодействие и пишем спектры взаимодействия через 10 мин. и через 50 мин. после начала взаимодействия. На глаз видно, что интенсивность падает. Постепенно интенсивность падает, потом, когда мы эти объекты снова разнимаем и записываем их по отдельности, то вы видите, что этот объект из такого спектра превратился в такой, такого – в такой, т.е. в обоих случаях интенсивность спектров упала. В общем, это согласуется с прежними данными.
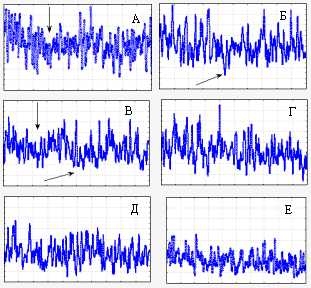
Рис. 7.. Изменения спектров Фурье в результате 1-часовых оптических взаимодействий между группами зародышей 14 и 31 стадий развития. Записи соответствуют 10-минутным отрезкам времени. Время накопления фотонов 0,1 с. А, Б: исходные спектры групп 14 и 31 стадий, соответственно. В, Г: их совместные спектры на 0-10 и на 50-60 минутах оптических взаимодействий, соответственно. Д, Е: спектры групп 14 и 31 стадий соответственно после окончания 1-часовых оптических взаимодействий. Обратите внимание на понижение спектральной плотности. Здесь и на Рис. 8 горизонтальная шкала охватывает диапазон частот от 3х10-3 Гц до 5 Гц.
Теперь я перехожу к результатам длительных оптических взаимодействий между зародышами. Приводятся в оптический контакт две партии зародышей. Оптический контакт, скажем, длится 12 ч. Затем берутся две партии зародышей из той же кладки, которые не находились в оптическом контакте и развивались те же самые 12 ч.
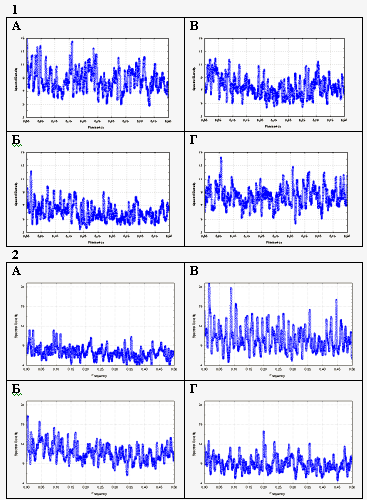
Рис. 8. . Спектры Фурье двух «четверок» образцов (1 и 2) до и после их длительного оптического взаимодействия. 1: взаимодействие зигот с зародышами на стадии бластулы. 2: взаимодействие стадии 2 бластомер с зародышами стадий 30-31 (перед вылуплением). А, В – спектры контрольных ранней и поздней групп зародышей соответственно. Б, Г – спектры тех же групп после оптического взаимодействия. В случае 1 спектр Б становится похожим на В (спад спектральной плотности в левой части), а спектр Г – на А (общий подъем спектральной плотности). В случае 2 спектр Б приближается к В (подъем спектральной плотности), а спектр Г – к А (падение спектральной плотности).
На Рис. 8 представлены спектры Фурье для двух разных пар (1 и 2) взаимодействующих партнеров разных стадий развития. При этом два верхних спектра от каждой пары (А и В) относятся к оптически не взаимодействовавшим партнерам, а нижние спектры (Б и Г) – к тем же партнерам после их 12-часового оптического взаимодействия. То есть в результате оптического взаимодействия спектр А трансформировался в Б, а спектр В – в Г. Подчеркнем, что стадии развития взаимодействовавших и не взаимодействовавших партнеров были, конечно, одни и те же. Что мы видим в результате? На спектре 1А амплитуды были выше, нежели на 1В, но после взаимодействия произошла инверсия – на 1Б амплитуды ниже, чем на 1Г. Аналогичная инверсия произошла на партнерах 2. Аналогичный «обмен амплитудами» мы видим во многих других случаях , но я сейчас сразу же перейду к тому, как мы количественно решили оценивать эти эффекты. Можно составить автокоррелограммы для каждого из этих спектров. Тогда мы получим очень загрубленную картину, но она позволит нам ввести некоторые количественные характеристики (Рис. 9)
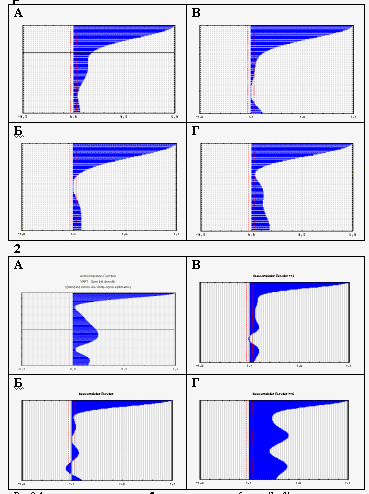
Рис. 9. . Автокореллограммы спектров Фурье двух «четверок» образцов (1 и 2) до и после их длительного оптического взаимодействия. 1: взаимодействие зигот с вылупившимися предличинками. Период накопления фотонов – 0,1 с. Максимальный лаг – 100 шагов (около 0,25 полного спектрального диапазона). 2: взаимодействие зигот с зародышами стадии 14. Период накопления фотонов – 0,5 с. Максимальный лаг – 200 шагов (около 0,5 полного спектрального диапазона). А, В – автокореллограммы ранней и поздней групп зародышей соответственно. Б, Г – автокореллограммы тех же групп после оптического взаимодействия. Отчетливо виден «обмен» характеристиками (Г становится похожим на А, Б – на В).
Мы берем здесь достаточно большое число лагов - более 100. Понятно, что значения коэффициентов корреляции всегда падают, потому что ближние значения всегда сильнее коррелируют, нежели дальние, но для нас интересен вот этот более дальний участок. Так вот: до взаимодействия этот объект имел такую автокоррелограмму , а второй объект имел такую автокоррелограмму . После взаимодействия получилось следующее: этот приобрел такую, а этот приобрел иную. Мы отсекаем ту часть автокоррелограммы, которая одинаковая во всех случаях – это примерно 30 ближайших значений, они имеют всегда высокий коэффициент автокорреляций, но они нам не интересны, и мы берем сумму коэффициентов корреляции в более дальней области, как бы интеграл от этой области. Мы видим, что у нас имеется весьма четкий обмен характеристиками вот в каком смысле. Если здесь интеграл, то есть сумма коэффициентов корреляции была выше, чем здесь, то послевзаимодействия там, где было больше – стало меньше, и наоборот.
Можно тут навести простенькую статистику. Обозначим оптически не взаимодействовавших (контрольных) партнеров как А1 и В1, а оптически взаимодействовавшие образцы из тех же кладок - А2 и В2. Будем оценивать влияние оптических взаимодействий на их спектры, определяя разности SС(ас) между образцами А2 и А1 (А2 – А1) и между образцами В2 и В1 (В2 – В1). Такие сравнения были произведены для 38 пар образцов, входящих в 19 «четверок» (А1, А2, В1, В2). Кроме того, для каждой четверки определяли величину
R = SС(ас)(A2) - SС(ас)(B2) / SС(ас)(B1) - SС(ас)(A1)
Очевидно, что R<-1 соответствует усилению (обострению) исходных различий между спектрами в результате оптических взаимодействий; при -1R>0 происходит инверсия свойств спектров, а при R>1 происходит инверсия с «перехлестом», то есть в результате оптических взаимодействий образцы не только «меняются» характеристиками своих спектров, но вновь приобретенные инвертированные различия оказываются сильнее исходных.
Как видно из Табл. 2, относящейся к длительным оптическим взаимодействиям, в 33 из 38 измеренных пар изменения SС(ас) каждого из образцов были направлены «навстречу» его партнеру (то есть у партнера с исходно меньшим значением SС(ас) оно увеличивалось, и обратно). Только в оставшихся 5 парах один из компонентов (и никогда – оба сразу) проявлял противоположное поведение (выделены жирным шрифтом в столбцах 4, 5 слева на Табл. 1). Измерения R показали, что у 14 «четверок» из общего числа 19 после оптических взаимодействий происходит инверсия значений SС(ас), причем у 11 из них – инверсия с «перехлестом». Из оставшихся 5 «четверок» у 4 наблюдается сглаживание различий по SС(ас), и лишь в одном случае – усиление исходных различий. Все эти различия по вышеупомянутому «критерию знаков» высоко достоверны. С другой стороны, из 8 «четверок», которые взаимодействовали в течение всего 60-100 минут, лишь в 4 случаях наблюдалась инверсия (правда, в одном из них– с «перехлестом»), в одном – сглаживание, а в трех других – даже некоторое усиление исходных различий между спектрами (Табл. 3). Таким образом, только длительные оптические взаимодействия (необходимые для возникновения аномалий развития) эффективно «обменивают» спектральные характеристики партнеров.
При группировке данных по возрастным группам (Табл. 2, два правые столбца) оказалось, что наибольшая инверсия (наивысшие значения R) после длительных оптических взаимодействий наблюдается в тех случаях, когда одним из партнеров являются зародыши средних стадий развития (бластула – гаструла), особенно если они взаимодействуют с зиготами. С другой стороны, если одним из партнеров являются предличинки (стадии 30-34) – инверсии наименьшие. Но это не означает, что SС(ас) в наибольшей степени изменяются обязательно у зародышей ранних или средних, а не старших стадий: окончательные величины инверсий являются функцией взаимодействующих пар, а не индивидуальным свойством отдельной популяции зародышей. Подчеркнем также отсутствие какой-либо связи между возрастом партнера и величиной SС(ас): относительно высокие значения данного показателя может иметь в равной степени как старший, так и младший оптический партнер.
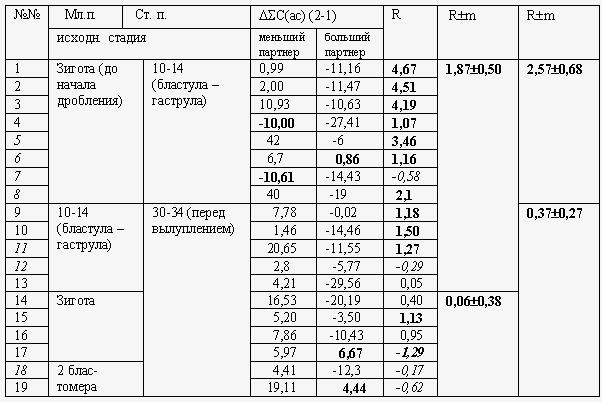
Табл. 2. Различия в автокорреляциях спектров Фурье образцов зародышей вьюна после 12-22-часовых оптических взаимодействий. В столбце « №№» курсивом обозначены измерения с периодом накопления фотонов 0,5 с. В остальных случаях период накопления 0,1 с. В столбцах «мл.п.» и «ст.п.» указаны и кратко описаны стадии развития (по Костомаровой, 1975), на которых началось взаимодействие младшего и старшего партнеров. В двух следующих столбцах жирным выделены случаи, нарушающие общую закономерность движения SС(ас) данного партнера навстречу другому. В графе «R» жирным выделены случаи инверсии с перехлестом, курсивом – случай сглаживания без инверсии, жирным курсивом – случай обострения исходных различий. Остальные случаи – инверсия без перехлеста. Два правые столбца – средние значения R ± ошибка разности для соответствующих групп данных
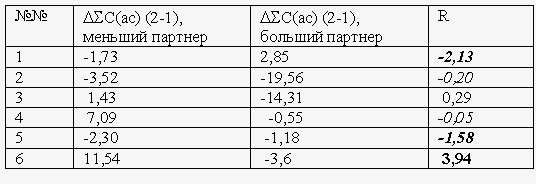
Табл. 3. Различия в автокорреляции спектров Фурье зародышей вьюна после 1-часовых оптических взаимодействий. Обозначения в столбце R как на Табл. 2
Это в сущности это все, пожалуй, что я хотел бы сказать по поводу этой группы опытов, которые связаны с взаимодействием зародышей и яйцеклеток вьюна.
Вывод такой: единственные характеристики, которые тут можно усмотреть, особенно для длительных взаимодействий, это взаимодействия частотно – амплитудного характера. Впечатление такое, что каждый из взаимодействующих зародышей воспринимает некий частотный спектр от другого объекта и перерабатывает внутри себя так, что происходит как бы обмен частотно – амплитудными характеристиками. Может быть здесь присутствует кто-нибудь, кто может указать на какие-нибудь другие случаи такого обмена, такого взаимодействия? Я достаточно случайно нашел работу, которая посвящена взаимодействию двух солитонов (Курин и Пермитин, 1987). В ней есть указание на то, что когда взаимодействуют два солитона, то они обмениваются между собой характеристиками. Вот такой сильно нелинейный эффект по-видимому здесь присутствует , что пожалуй, может объяснить наблюдаемое явление.
Теперь в оставшейся части моего сообщения я хотел бы поговорить о таких вещах, когда взаимодействуют два объекта, один из которых является чисто биологическим, а другой не совсем биологическим или совсем небиологическим.
Я поясню, о чем идет речь. Очень интересные взаимодействия существуют, например, между самими зародышами и их оболочками (Белоусов с соавт., 1997). Вот скажем в курином яйце между зародышем и скорлупой. Дело в том, что куриное яйцо, просто если его поместить в камеру ФЭУ – оно достаточно сильно светит. Этот сигнал плавно падает в темноте, и в какой-то степени зависит от предыдущей засветки, но и изолированная скорлупа куриного яйца, из которого удалено содержимое, довольно сильно светит. Если изолировать ранний зародыш сам по себе - этот крохотный бластодиск, то он дает излучение порядка 15 импульсов в секунду, если этот зародыш держать на желтке, вместе с желтком, то он дает сигнал порядка 50 импульсов в секунду. А вот скажем, если взять целое яйцо или его скорлупу, они дают значительно больший сигнал - порядка 2500 импульсов в секунду. Если мы возьмем зародыш более поздних стадий развития, то удивительным образом сам зародыш вообще не дает никакого свечения, а зато скорлупа дает излучение порядка 5000 импульсов в секунду. Оказалось, что между этими двумя компонентами, между зародышем и скорлупой, существуют достаточно необычные взаимодействия. Можно изготовить такой препарат: аккуратно вылить желток с зародышем в кювету и накрыть его куском скорлупы от этого яйца и поставить ФЭУ мерить его сигнал. Окажется следующее (Рис. 10).
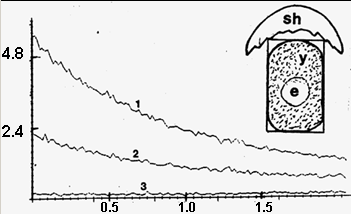
Рис 10/ Записи сверхслабых излучений от помещенного в кварцевую кювету желтка 2-дневного развивающегося куриного яйца (y) с бластодермой (е). Кювета накрыта кусочком скорлупы от того же яйца (sh) и обращена бластодермой к катоду фотоэлектронного умножителя. 1 – запись излучения от желтка + кусочек скорлупы. 2 – запись излучения от того же кусочка скорлупы без желтка. 3 – запись излучения от одного желтка без скорлупы. Видно, что излучение 1 существенно выше суммы 2 и 3. Горизонтальная ось: минуты. Вертикальная ось: количество импульсов за 1с х 1000
Если вы будете измерять сигнал от всей этой системы, то получится довольно высокий, хотя и плавно падающий сигнал (кривая 1). Теперь, если желтка нет, вы этот самый кусочек скорлупы ставите отдельно и от него меряете значение сигнала, то его значение значительно меньше – это кривая 2. Но отсюда никак не следует, что 1 есть простая сумма излучений желтка и скорлупы, потому что если вы будете мерить излучение только от одного желтка, то получите кривую 3. Сумма сигналов 3+2 явно меньше сигнала, показанного кривой 1, так что здесь имеется несомненное неаддитивное взаимодействие. В данном случае желток как бы « стимулирует» излучение скорлупы. Но может быть и обратное. Это зависит от стадии развития зародыша и вообще от того, развивается ли зародыш. Обнаружено (Белоусов с соавт., 1997), что лишь у неоплодотворенных яиц (которые мы употребляем в пищу) излучение целого яйца практически равно излучению его скорлупы. Между тем, у яиц примерно двух дней развития (подобно выше рассмотренному) целое яйцо светит сильнее одной лишь его скорлупы, а у яйца девяти дней развития соотношение обратное: целое яйцо светит по большей части слабее одной лишь его скорлупы. Отсюда можно сделать выводы, что на ранних стадиях развития происходит некое «закачивание» излучения в скорлупу, а на более поздних стадиях наоборот - «выкачивание» фотонов из скорлупы. В любом случае мы имеем нарушение чисто аддитивных соотношений.
То же самое наблюдается в яйцеклетках амфибий. Яйцеклетки амфибий одеты слизистой оболочкой, я думаю, что те, кто когда-нибудь видели лягушиную икру обращал внимание: она особенно хорошо светится в лучах солнца и действительно, наверное, принимает участие в восприятии и переработки света.
Так вот, если взять относительно раннюю стадию развития (стадию гаструлы) и сравнить интенсивность излучения порции яйцеклеток, одетых оболочками, с излучением одних лишь снятых с них оболочек, то получится, что последние светят слабее. Это, конечно, не вызывает удивления. Но если взять зародышей более поздних стадий развития (нейрула), то эти соотношения инвертируются так же, как у куриных яиц. А именно, оказывается, что изолированные оболочки светят больше, чем когда они одевали зародыши (Белоусов и Лучинская, 1998). Вывод можно сделать тот же зародыши как бы «подсасывают» некоторое количество фотонов из оболочек, отчего они и светят меньше. Вот такие соотношения имеют место, их довольно много и может быть это даже специальная тема для разговора. В частности, я знаю, что есть аналогичные данные на образцах крови (Воейков и Новиков, 1997).
В самом конце я хотел бы очень коротко сказать о некоторых данных по взаимодействию яйцеклеток с совсем уже неживыми источниками излучений. Существует такой источник сверхслабого излучения, который был открыт еще в лаборатории Гурвича. Если смешать аминокислоту – глицин – с перекисью водорода, то наблюдается довольно сильная вспышка излучения, интенсивность которой через некоторое время падает.
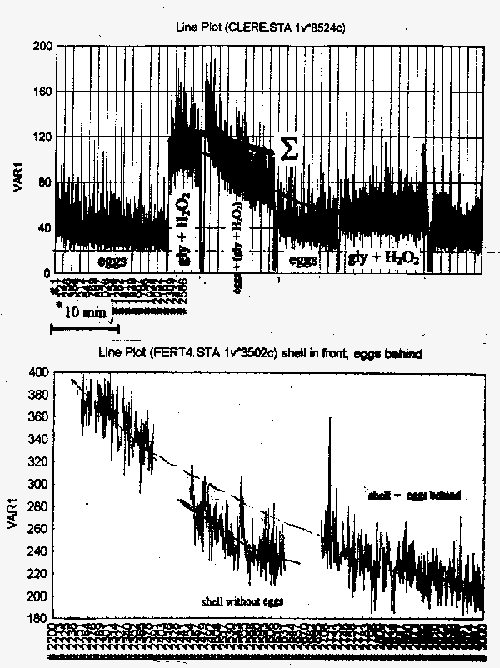
Рис 11 Взаимодействия развивающихся яйцеклеток амфибий с неорганическими источниками сверхслабых излучений. А: взаимодействия яйцеклеток с реакцией окисления глицина перекисью водорода. Б: взаимодействия яйцеклеток с фрагментом скорлупы неоплодотворенного куриного яйца. Горизонтальная ось: время (указан 10-минутный промежуток), вертикальная ось – количество импульсов за 1 с. Объяснения в тексте
В данном случае бралась кювета с двумя химически изолированными отделениями, разгороженными кварцем, в одно отделение помещались развивающиеся яйца, в другом отделении осуществлялась реакция глицина с перекисью водорода. Сначала записываем излучение от яиц (рис. 11А, eggs), потом мы помещаем сюда глицин с перекисью водорода , записываем сигнал только от него (рис. 11А, gly + H2O2) – вот он такой большой, а потом мы записываем совместный сигнал этой реакции вместе с яйцами (Рис. 11А, eggs + (gly + H2O2)). Мы наблюдаем, что в первый момент происходит очень сильный выброс свечения, который затем очень быстро падает вниз. Выброс превышает интенсивность свечения самой по себе реакции. Очевидно, что яйца, которые сами по себе светят достаточно мало, вступают в некие взаимодействия с этим химическим источником и дают такую вспышку, которая затем падает. Интересно, что так же можно заставить оптически взаимодействовать развивающиеся яйцеклетки амфибий со скорлупой куриного яйца, казалось бы, совершенно чуждым для них объектом (Рис. 11Б). Вот так светит скорлупа куриного яйца в том случае, когда сзади от нее находятся яйцеклетки амфибий (левая часть кривой). Если яйцеклетки убрать (shell without eggs), то скорлупа светит существенно меньше, на более низком уровне. Если мы снова ставим яйцеклетки позади скорлупы (shell + eggs behind), то излучение поднимается на значительно большую величину, нежели мы могли бы ожидать от одних только яйцеклеток – особенно если учесть, что «формально» скорлупа непроницаема для столь малых световых потоков. Снова мы имеем неаддитивное взаимодействие.
Ну и в сущности, пожалуй, я на этом кончу. Наверно, мой доклад и не должен содержать каких-либо выводов, Я просто хотел вам рассказать о том какой это странный мир сверхслабых излучений и как тут может происходить взаимодействие биологических и не биологических объектов. Спасибо за внимание. [Более подробное изложение приведенных в докладе данных содержится в статье Белоусова с соавт., 2003].
Владимирский Б.М.: кто желает получить дополнительную информацию из этого странного мира?
Вопрос: почему всегда вы не делали опыт отдельно, контроль отдельно, контроль, потом на том же месте опыт. С течением времени объекты развиваются, они же живые, разорваны на кусочки и нельзя выделить эксперимент, сравнить его две системы с одним и тем же, этапы, контроль и опыт отдельно
Ответ: у меня это было и даже было показано при взаимодействии , скажем, некая стадия, я приводу ее во взаимодействие, потом я пишу ее отдельно и т.д.
Вопрос: вы сравниваете всегда два объекта до взаимодействия, потом во время взаимодействия, потом без взаимодействия. С самого края нужно было еще две колонки поместить, что делают из одной партии и из другой партии образца. Убранные вообще в другую сторону, как они ведут себя? Вы это знаете, вы возможно смотрели как они обычно ведут, вы это уже знаете, не возникает никакого вопроса в этом, но для всякого человека, который сморит первый раз, напрашивается такая вещь: у вас всюду две колонки, образец один и образец другой, до взаимодействия, потом они взаимодействуют, оказывается примерно такая схема демонстрации А что, а как ведет себя образец аналогичный правой кювете и аналогичный левой кювете вне взаимодействия, в течение времени. Может быть они от времени так меняются?
Ответ: нет
Брунс: я понимаю, что нет, у вас уже опыт и вы знаете как они себя ведут, но когда смотрит посторонний человек, хотелось бы, чтобы образец правой кюветы и образец левой кюветы без взаимодействия то же показывались: как они ведут себя со временем. Время тут фактор.
Ответ: вопрос ясен. Везде, где я говорил о результатах длительных взаимодействий у меня были 4 картинки. Две из них демонстрируют образцы, которые не взаимодействовали между собой. Другие две картинки – это образцы, которые взаимодействовали между собой. Вот которые не взаимодействовали и которые взаимодействовали – они абсолютно одной стадии развития Т.е. во всех этих опытах я сравниваю одинаковые стадии развития, это раз. Теперь ваш второй вопрос – меняется ли что нибудь без взаимодействия в течение тех 12 часов, что объекты взаимодействуют? – Конечно, меняется, об этом было в начале доклада. Я говорил, что по ходу развития спектры Фурье меняются.
Вопрос: сколько времени облучается объект взаимодействия?
Ответ: сколько времени ведется запись? Я говорил, что запись ведется 10 минут, за это время никаких существенных различий по развитию не происходит – это раз. Причем каждый раз сравниваются одинаковые стадии развития. Скажем, если вы берете 12 часовые периоды, то сравниваются взаимодействия из одних и тех же кладок и из одних и тех же стадий развития. То же самое относится и к ранним стадиям развития объекта
Вопрос: Во-первых, по классическому опыту самому первому, с оптическим взаимодействием препаратов, вы сказали, что кварц не разрывал взаимодействия, а стекло разрывает, в то же время это кюветы с прозрачным дном?
Ответ: да
Вопрос: а если (неразборчиво) то каково поглощение воды, если там вода, то это нужно проконтролировать
Ответ: Там верхний образец просто лежит на дне, под ним воды нет, нижний образец, там по данным Бурлакова должно быть не более 2 мм воды на ним.
Вопрос: я хотел бы спросить вас, вроде бы аудитория уже устала спрашивать. Это что, нужно рассматривать, вы о теории ничего не говорили, можно сказать по крайней мере несколько слов для тех, кто далеко стоит от этих исследований. Это какое-то кодовое, по-видимому, воздействие и вот для того, чтобы разобраться в этом надо, собственно говоря, расшифровать этот код.
Ответ: здесь существуют разные аспекты. Вот это та идеология, которой Фриз Попп придерживается, с которым мы работаем вместе, это такое нелинейное взаимодействие сближенных осцилляторов, существует некая теория Дикке. Согласно этой теории, если устанавливается взаимодействие, то очень часто возникает режим, который называется субрадианс - субрадиация, т.е., скажем, совокупный сигнал понижает свою интенсивность по сравнению с каждым сигналами по–отдельности. Происходит нечто вроде интерференции. У него такие представления. И в этом смысле те результаты, когда при кратковременном взаимодействии общий сигнал падает, как будто укладываются в эти представления о субрадиансе. Но теперь, если говорить о более длительных взаимодействиях, которые дают морфологические эффекты , по-видимому здесь надо говорить о том, что каждая партия зародышей, если они разных стадий, имеет свой собственный частотный спектр и если в него вмешаться другим частотным спектрам, то для зародышей это плохо. Причем особенно важен этот частотный спектр для ранних стадий развития. Потому что на ранних стадиях развития он приурочен к клеточным циклам, я и на прошлых докладах, и осенью об этом говорил , просто не стал на этом сейчас останавливаться [см. Белоусов с соавт., 2002]. Там очень четко видно, что на ранних стадиях развития каждый клеточный цикл начинается с серии вспышек, все идет в определенной ритмике. Если навязать образцам какую-то другую ритмику , то это вносит сбой. Вот что можно сегодня об этом сказать.
Владимирский: Еще один вопрос такой, наверное, для тех, кто далек от этих исследований Вы ограничились диапазонами длин волн от короткого ультрафиолета до 800 нм . А в инфракрасной области, например, могли бы быть обнаружены такие эффекты?
Ответ: вероятно. Это очень интересные области, но просто приборная база в лабораториях плохо позволяет это выявить. Я знаю, что с инфракрасными измерениями тут большие трудности. Вероятно, сюда простираются эти эффекты.
Владимирский: Спасибо!
Вопрос: если с биологической точки зрения рассматривать, может это действительно некий гипотетический механизм, который заложен в естественном отборе. Например, когда откладывается икра рыбами на разных стадиях, и если это происходит чуть раньше, то они имеют шанс больше своего потомства вывести в люди, по сравнению с теми, которые пришли на то же самое место позже и отложили икру позже , то у них шанс меньше вывести здоровое потомство
Ответ: Да, Бурлаков – ихтиолог, он начал с того, что обратил внимание на то, что рыба никогда не отложит икру на уже отложенную икру, как будто бы знает об этих взаимодействиях . Но я думаю, что это не главное . Наиболее важны синхронизирующие сигналы внутри кладки. Вот они наиболее важны. Давно известны групповые эффекты, что икринки вместе развиваются лучше, чем по отдельности. Нужно здесь разделить оптические и химические взаимодействия. И вот такие опыты показывают, что оптический канал имеет место.
Вопрос: Если поставить такой опыт : есть один образец, есть второй, здесь происходило оптическое взаимодействие живой объект – живой объект. Насколько он промодулирован, скажем, информационным каким-то потокам….(неразборчиво). Если сделать такой опыт: взять такой же спектр частот от биологического объекта, записать его и его же излучить с теми же интенсивностью от другого источника. И потом , на основании этого сделать выводы: есть ли помимо интенсивности этого информационная компонента, которая излучается в воде
Ответ: Бурлаков делал подобные опыты. Они дали положительный результат: он взял записи сверхслабых излучений и промодулировал их в инфракрасной области с помощью лазера и получил результаты, соответствющие взаимодействиям живых объектов.
Конец пленки.
|