|
Вверх
на главную страницу
История
Факты
Идеи
Люди
Библиография
Конференции
В.В. Леднев Биологические эффекты крайне слабых магнитных полей: идентификация первичных мишеней |
|
В.В. Леднев, Н.А. Белова, З.Е. Рождественская, Х.П. Тирас Представлены результаты исследований, свидетельствующие о том, что крайне слабые переменные магнитные поля (КС ПеМП) с амплитудами в области «пикотеслового», «нанотеслового» и «микротеслового» диапазонов оказывают существенное влияние на биологические и физико-химические процессы, а именно: регенерацию плоских червей (планарий Dugesia tigrina), гравитропизм в отрезках стеблей льна, а также на кристаллизацию карбоната кальция (СаСО3) из солевых растворов. Теоретический анализ полученных данных позволяет сделать вывод о том, что первичным рецептором КС ПеМП в биосистемах является водно-солевая компонента. Сопоставление полученных результатов с данными о геомагнитных предвестниках землетрясений позволяет рассматривать влияние на биообъекты изменений в частотно-амплитудном спектре геомагнитных пульсаций, происходящих перед землетрясениями, как весьма вероятный механизм биологических предвестников землетрясений.
Введение Как известно, землетрясения часто предваряются различными геофизическими и геохимическими предвестниками, включая усиление микросейсмической активности и акустических шумов, выхода газов из грунта (например, радиоактивного радона), повышение концентрации аэроинов [Сидорин, 1992]. При этом каждый из перечисленных факторов по отдельности или в сочетании друг с другом может рассматриваться как возможный стимул необычного поведения животных. Тем не менее, некоторые авторы полагают, что электромагнитные возмущения, предшествующие землетрясениям, более полно соответствует совокупности наблюдений, связанных с биопредвестниками [Красногорская и др., 1984; Темурьянц и др., 1992]. Очевидно, что получение полной картины изменений электромагнитного поля (ЭМП) в период, предшествующий землетрясениям, – исключительно сложная задача. Тем не менее, за последние десятилетия было выполнено достаточно большое число измерений в широкой области частот и амплитуд спектра электромагнитных колебаний, результаты которых показывают, что наиболее распространенным видом возмущений ЭМП является появление импульсов широкого спектра частот в диапазоне от долей герц до нескольких мегагерц, включая диапазоны регулярных геомагнитных пульсаций и шумановских резонансов [Темурьянц и др., 1992]. Однако амплитуда указанных возмущений ЭМП, как правило, очень мала – для переменных магнитных полей она соответствует единицам нанотесла, но, как правило, меньше, что, естественно, вызывает вопрос о возможности восприятия таких полей биологическими системами. К настоящему времени твердо установлено, что низкочастотные переменные магнитные поля с амплитудами (величинами магнитной индукции), равными или превышающими примерно 10 мкТл, способны индуцировать биологические эффекты в ряде биосистем. Вместе с тем, вопрос о возможности воздействия на человека, животных и растений значительно более слабых или крайне слабых переменных магнитных полей (КС ПеМП) с амплитудами меньше 10 мкТл и, в особенности, интересующими нас в данном случае полей «нанотеслового» и «пикотеслового» диапазонов, является предметом острых дискуссий. С одной стороны, появляется все больше экспериментальных доказательств того, что КС ПеМП «нанотеслового» диапазона способны оказывать существенное воздействие на ряд биологических процессов (перечень работ приведен в [Темурьянц и др., 1992; Белова, 2001]). С другой стороны, в некоторых публикациях ставится под сомнение даже принципиальная возможность биологических эффектов КС ПеМП [Adair, 1991, 1999; Valberg et al., 1997]. Имеются лишь единичные сообщения о результатах лабораторных исследований возможной «биологической эффективности» КС ПеМП «пикотеслового» диапазона [Темурьянц, 1972; Белова, Леднев, 2001]. Очевидно, что развитие представлений относительно возможной причинной связи между биопредвестниками землетрясений и слабыми вариациями магнитных (электромагнитных) полей в существенной степени зависит от прогресса в исследованиях взаимодействия КС ПеМП с биосистемами. В данной работе представлены результаты исследований, свидетельствующие о том, что КС ПеМП с амплитудами в области «пикотеслового», «нанотеслового» и «микротеслового» диапазонов оказывают существенное влияние на биологические и физико-химические процессы, а именно, регенерацию плоских червей (планарий Dugesia tigrina), гравитропизм в отрезках стеблей льна, а также на кристаллизацию карбоната кальция (СаСО3) из солевых растворов. Теоретический анализ полученных данных позволяет идентифицировать первичные мишени воздействия КС ПеМП в биосистемах.
Методы исследований
Экспонирование всех изучавшихся тест-систем в КС ПеМП с величиной магнитной индукции BAC осуществляли на фоне постоянного магнитного поля Земли с величиной магнитной индукции (амплитудой) BDC= 42 мкТл. Соответственно, в общем случае термин КС ПеМП фактически означает, что используется комбинированное (модулированное) магнитное поле
Использовались КС ПеМП со следующими параметрами: Величина фонового переменного магнитного поля на частоте 50 Гц в месте постановки экспериментов составляла 15 нТл. Все опыты проводили в интервале температур 18–20°°С. Тест-система 1: регенерирующие плоские черви (планарии Dugesia tigrina).
Исследуемый биопроцесс: скорость регенерации ампутированной головной части планарий.
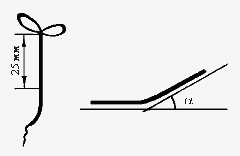
Изображения регенерирующих планарий вводили в компьютер с помощью видеокамеры. Используя специальную программу, определяли площадь S всей планарии и площадь s прозрачной регенерирующей бластемы (рис.1). Результаты этих измерений использовали для вычисления среднего (по 30 регенерантам) значения индекса регенерации R=s/S для контрольных и опытных животных.  где dэ и dk – стандартные ошибки cредних величин. 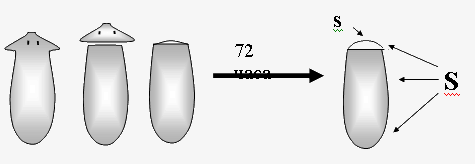 Рис. 1. Схема измерения индекса регенерации планарий. s – площадь вновь образовавшейся ткани (бластемы); S – площадь всего тела планарии; s/S – индекс регенерации Тест-система 2. Сегменты стеблей проростков льна. Исследуемый биопроцесс: гравитропический изгиб апикальных участков сегментов льна, сопровождающий их поворот из вертикального в горизонтальное положение. Метод регистрации эффекта КС ПеМП: измерение среднего угла изгиба апикального конца сегмента относительно горизонтальной плоскости. Подготовка сегментов растений: Проростки льна (Linum bienne) выращивали из семян в термостате при 26°°С в полной темноте в течение 3–5 сут в специальных растильнях. Через 3–4 сут стебли льна достигали длины 3–4 см. От верхушек стебля льна отрезали листья так, что отрезки содержали только апикальную часть стебля длиной 25 мм. Стимуляция гравитропического ответа. Отрезки стеблей льна раскладывали в чашки Петри диаметром 90 мм на фильтровальной бумаге, смоченной 2 мл дистиллированной воды, по 20 отрезков на чашку. Положение базальных концов отрезков фиксировали, накладывая на них силиконовые кольца. Как в «опыт», так и в «контроль» ставили по 2 чашки Петри. Примерно через 30 мин горизонтально расположенные отрезки начинали изгибаться, так что через 2–3 ч угол изгиба составлял несколько десятков градусов.
Величину гравитропического ответа определяли путем измерения среднего по числу отрезков угла изгиба (отклонения конца отрезка от горизонтальной плоскости, (рис.2)) , где d – стандартная ошибка средней величины. Измерения проводили через 2–3 ч после начала опыта с помощью транспортира. Величину эффекта МП выражали как относительную разницу (в %) между средними углами aэ и ak гравитропического изгиба сегментов, измеренных, соответственно, в опытных, т.е. экспонированных в КС ПеМП, и в контрольных образцах, согласно выражению 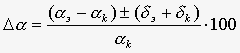 где dэ и dk – стандартные ошибки средних величин углов изгиба.
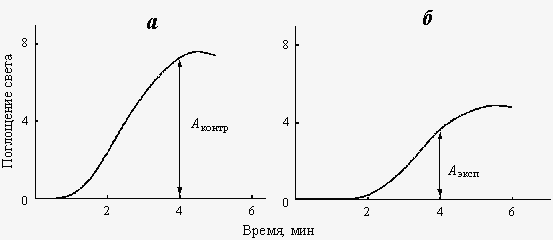 Рис. 3. Типичные кривые изменения поглощения света при кристаллизации СаСО3 из контрольных (а) и экспонированных в магнитном поле (б) растворов. Аконтр и Аэксп – поглощение света в определенный момент времени после начала кристаллизации. Тест-система 3: солевые растворы хлористого кальция (СаСl2) и карбоната натрия (Na2CO3). Исследуемый физико-химический процесс: скорость кристаллизации карбоната кальция СаCO3, образующегося при смешивании растворов солей СаСl2 и Na2CO3. Метод регистрации эффекта КС ПеМП: измерение динамики светопоглощения в растворе с помощью фотоэлектрического калориметра. Протокол эксперимента. Приготавливали растворы СаСl2 (6 мМ) и Na2CO3 (12 мМ) в дистиллированной воде. Растворы разливали в стеклянные стаканчики объемом 100 мл, по 90 мл в каждый. Опытные растворы СаСl2 и Na2CO3 экспонировали в соответствующем КС ПеМП, в то время как такие же контрольные растворы находились в магнитном поле Земли. Время экспонирования составляло от 6 до 12 ч. Реакцию кристаллизации СаCO3 инициировали смешиванием равных объемов (по 4 мл) растворов СаСl2 и Na2CO3 в стеклянных кюветах, с длиной оптического пути 2 см. Измерение динамики светопоглощения в процессе кристаллизации СаСО3 (рис.3) проводили в фотоэлектрическом калориметре КФК-2 на длине волны 590 нм, регистрируя сигнал на самописце LKB 2210.
Результаты
Важный результат экспериментальных исследований состоит в том, что биоэффекты «нанотесловых» и «микротесловых» полей проявляются лишь при определенных соотношениях амплитуды и частоты поля (рис.5). В частности, в поле с амплитудой 134 нТл, биоэффекты поля наблюдаются на частотах 3 и 5 Гц, в то время как на частоте 4 Гц эффект отсутствует (рис.5,а). При одновременном увеличении как амплитуды так и частоты поля в 3 раза (BAC=402 нТл, fAC=9, 12, 15 Гц), характер зависимости величины биоэффекта от частоты поля не изменяется – эффект поля наблюдается на частотах 9 и 15 Гц и отсутствует на частоте 12 Гц (рис.5,б). При дальнейшем увеличении амплитуды поля до «микротесловых» значений (BAC=1608 нТл) «двугорбый» характер зависимости величины биоэффекта от частоты поля сохраняется (рис.5,в), однако, в этом случае наблюдается сдвиг положения минимума («нулевого» эффекта) от значения BAC/fAC=33.5 (при амплитудах 134–402 нТл) до значения BAC/fAC=44.7 (при амплитуде 1608 нТл). Также воздействие «нанотеслового» поля (BAC=134 нТл, fAC=3, 4, 5 Гц) на угол гравитропического изгиба в сегментах стеблей льна, сопровождается увеличением гравитропической реакции на частотах 3 и 5 Гц, при отсутствии биоэффекта на частоте 4 Гц (рис.6).
Экспонирование растворов СаСl2 и Na2CO3 в «пикотесловых» и «нанотесловых» полях приводит к снижению скорости кристаллизации СаCO3, происходящем при их последующем смешивании. Соответствующие результаты приведены на рис.7. 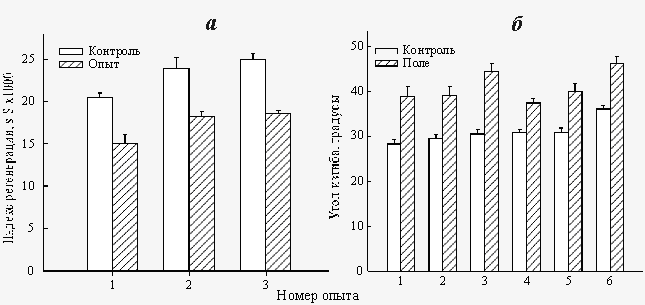 Рис. 4. Влияние «пикотеслового» КС ПеМП (BAC=640 пТл, fAC=10 Гц) на скорость регенерации планарий (а) и на гравитропический изгиб в сегментах стеблей льна (б)
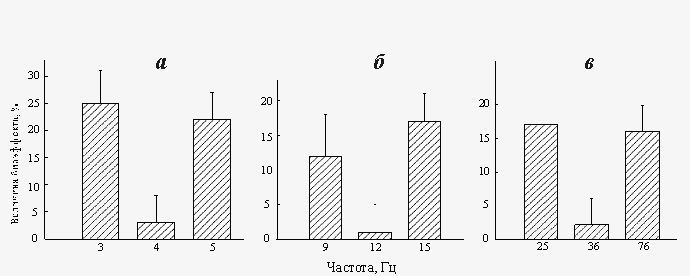 Рис. 5. Зависимость скорости регенерации планарий от параметров от «нанотеслового» и «микротеслового» КС ПеМП BAC=134 нТл, fAC=3, 4 и 5 Гц (а); BAC=402 нТл, fAC=9, 12 и 15 Гц (б); BAC=1608 нТл, fAC=25, 36 и 76 Гц (в)
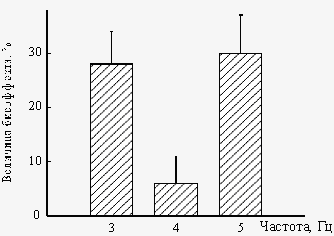 Рис. 6. Зависимость гравитропического изгиба в сегментах стеблей льна от частоты «нанотеслового» КС ПеМП (BAC=134 нТл, fAC=3, 4 и 5 Гц)
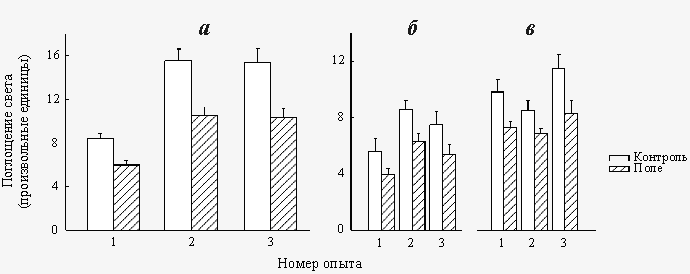 Рис. 7. Влияние предварительного экспонирования растворов CaCl2 и Na2CO3 в «пикотесловом» (BAC=640 пТл, fAC=10 Гц – а), в «нанотесловом» (BAC=134 нТл, fAC=5.0 Гц – б) и (BAC=134 нТл, fAC=6.3 Гц – в) КС ПеМП на скорость кристаллизации СаСО3 Обсуждение результатов Экспериментальные данные, представленные на рис.4–6, свидетельствуют о том, что КС ПеМП с величинами магнитной индукции в области «пикотеслового» и «нанотеслового» диапазонов способны оказывать существенное воздействие на биосистемы как животного, так и растительного происхождения. Биологическая эффективность КС ПеМП «нанотеслового» диапазона была показана также рядом авторов (см. обзоры в [Темурьянц и др., 1992; Белова, 2001]), в то время как биоэффекты «пикотесловых» синусоидальных магнитных полей обнаружена нами впервые. Способность КС ПеМП воздействовать на биосистемы при амплитуде поля всего лишь 640 пТл означает необходимость пересмотра широко распространенного представления о том, что минимальная, или «пороговая», амплитуда переменного магнитного поля, обеспечивающая возможность индукции биологических эффектов, составляет несколько единиц микротесла. Более того, вполне возможно, что «пороговые» величины магнитной индукции КС ПеМП могут оказаться намного ниже 640 пТл. При этом важно отметить, что биосистемы оказались способными детектировать «пикотесловые» магнитные поля на фоне значительно более интенсивного (примерно в 30 раз) магнитного шума промышленной частоты (50 Гц). Как следует из рис.7, «биологически эффективные» «пикотесловые» и «нанотесловые» поля оказывают также существенное влияние на свойства водно-солевых растворов СаСl2 и Na2CO3. Эти данные позволяют предположить, что воздействие КС ПеМП на биосистемы, по крайней мере, с теми параметрами, которые были использованы в настоящей работе, осуществляется через их водно-солевую компоненту. Наличие определенных закономерностей зависимости величины биоэффектов от соотношения амплитуда/частота, а именно: наличие резко выраженных «провалов» биологической эффективности КС ПеМП при определенных соотношениях параметра амплитуда/частота, а также сохранение биоэффектов при одновременном пропорциональном изменении амплитуды и частоты поля, позволяет сделать предположение о «первичных» мишенях «нанотесловых» и «пикотесловых» магнитных полей, которыми, по нашему мнению, являются соответственно магнитные моменты спинов ядер водорода и диамагнитного тока электронов в биомакромолекулах, а также в молекулах воды. Детальное рассмотрение этих вопросов выходит за рамки данной статьи. Отметим лишь, что в основе взаимодействия КС ПеМП и тест-систем различных типов лежит частотно-амплитудная модуляция электромагнитного поля, создаваемого вследствие прецессии соответствующих магнитных моментов в постоянном магнитном поле Земли. Возможность появления биологических и физико-химических эффектов КС ПеМП – это, в конечном счете, следствие важной особенности частотной модуляции, заключающееся в том, что амплитуды ближайших к несущей частоте сателлитов могут быть большими (при определенных соотношениях между амплитудой и частотой переменной компоненты поля) даже при очень малых величинах магнитной индукции переменной компоненты поля (КС ПеМП). Именно при этих условиях величина биологических эффектов КС ПеМП определяется интенсивностью боковых сателлитов, ближайших к несущей частоте. Совокупность результатов, представленных в данной работе, показывает, что между «аномальным» поведением ряда «биообъектов» и возмущением ЭМП, появляющимся перед землетрясениями, может существовать не только корреляция, но и причинно-следственная связь. В частности, параметры переменного магнитного поля, зарегистрированного перед сильными землетрясениями на Камчатке (амплитуда 0.5–1.0 нТл, частота 10 Гц) [Соболев и др., 1974], близко соответствуют «пикотесловому» КС ПеМП, биологическая эффективность которого была показана в данной работе. Заметим, однако, что возможность индукции «аномального» поведения животных перед землетрясениями крайне слабыми переменными магнитными полями не ограничивается схемой воздействия на биообъект комбинированного магнитного поля со слабой переменной компонентой. Теоретически возможна также ситуация, при которой подавляется повседневное («нормализующее», «синхронизирующее») биологическое действие переменных полей, например действие регулярных Рс колебаний геомагнитного поля или шумановских резонансов, в результате появления перед землетрясением магнитного шума в соответствующей полосе частот или резкого ослабления интенсивности указанных колебаний поля. Влияние переменных электрических полей с параметрами, имитирующими шумановские резонансы (амплитуда 1 мВ/м, частота 10 Гц) на психофизические показатели человека было обнаружено Х.Кёнигом вскоре после открытия этого типа глобальных электромагнитных колебаний [Konig, 1974]. Кроме того, возможная роль шумановских резонансов в качестве суточного задатчика времени у человека была показана в экспериментах Р.Вевера [Wever, 1974]. Имеются также десятки публикаций об успешном использовании переменных магнитных полей с амплитудой 6–7 пТл и частотой 7–8 Гц в терапии ряда заболеваний центральной нервной системы (см., например, [Sandyck, Derpapas, 1993; Sandyck, 1994; Jacobson, 1994]). Тем не менее, несмотря на несомненную привлекательность, гипотеза о возможности влияния магнитной компоненты шумановских резонансов на биосистемы до сих пор не была подвергнута серьезной экспериментальной проверке. По-видимому, основная причина отсутствия исследований в этой области, заключается в априорном отрицании возможной биологической эффективности сверхслабых магнитных полей, характерных для резонансов Шумана: (1–7 пТл на частоте 8 Гц [Williams, 1996]). Представленные в настоящей работе данные относительно биологических и физико-химических эффектов «пикотесловых» КС ПеМП позволяют считать, что исключение из рассмотрения в качестве возможного механизма биологических предвестников землетрясений влияния на биообъекты изменений в частотно-амплитудном спектре геомагнитного поля, происходящих перед землетрясениями, было бы явно преждевременным.
|
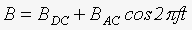 ,
,